A doksi online olvasásához kérlek jelentkezz be!
A doksi online olvasásához kérlek jelentkezz be!
Nincs még értékelés. Legyél Te az első!





Tartalmi kivonat
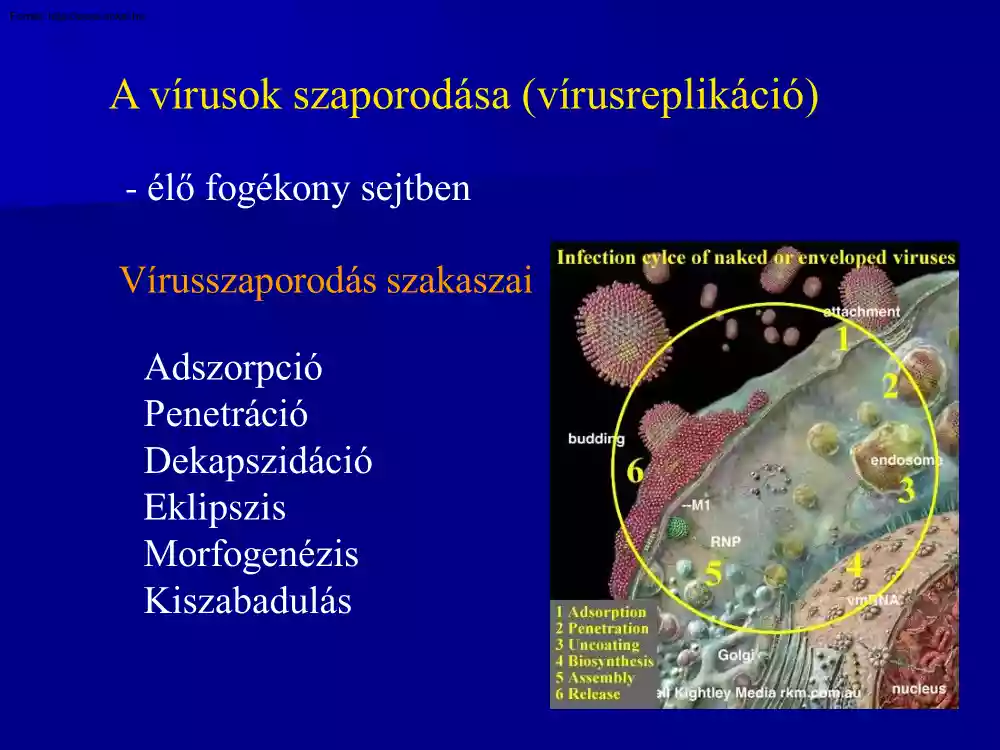
A vírusok szaporodása (vírusreplikáció) - élő fogékony sejtben Vírusszaporodás szakaszai Adszorpció Penetráció Dekapszidáció Eklipszis Morfogenézis Kiszabadulás 1. Megtapadás - Adszorbció a vírus - sejt kapcsolat első lépése. - a sejt membránján lévő receptorokhoz történik a vírus kötődése . - a receptorok szintézisét a sejtgenom határozza meg. Az a sejt illetve szervezet amely nem tartalmaz fajlagos receptorokat egy adott vírus számára ellenálló az illető vírussal szemben. - a növényi sejtfal nem tartalmaz vírus receptorokat, a növénypatogén vírusok megtapadásához előfeltétel a sejtfal mechanikai sérülése. - a humán- és állatpatogén vírusok kapszidján olyan szerkezetek találhatók melyek képesek kötődni a sejtek felszínén lévő receptorokhoz. - Ca és Mg ionok elősegítik a folyamatot. - a sejt típusa, kora, anyagcseréje, valamint környezete befolyásolja a víruskötőhelyek számát és
hozzáférhetőségét. - a baktériumok fágkötő receptorai a sejtfalban találhatók. - a Gram negatív baktériumok esetében lipopoliszaharid természetűek, a Gram pozitív baktériumoknál mureinhez kötött teichoinsavakból állnak. - a fágok a farok rostjaival és alaplapjával, illetve poliszaharidáz enzim segítségével valósítják meg a kötődést a baktériumsejt receptoraihoz. 2. Bejutás - penetráció - a növénypatogén vírusok esetében a teljes virion hatol be a növényi gazdasejtbe. A sejtfalak mechanikai sérülését követi, amit az ízeltlábúak szúrásai, jégverés, növényápolási munkálatok okoznak. - a humán- és állatpatogén vírusok behatolása 2 mechanizmus révén valósul meg: endocitózis sejtfúzió - endocitózis során a gazdasejt membránja betűremkedik és bekebelezi a viriont mely egy vakuólába jut. A vírus bekebelezése után helyreáll a sejthártya épsége. - a penetrációnak ezt a formáját
viropexisnek is nevezik. - fúzió során a vírusburok (peplon) fuzionál a sejtmembránnal, egy csatorna képződik amelyen át a vírus bejut a citoplazmába. - a bakterifágok esetében a kontraktilis fehérjetok összehúzódik, a fág belöveli a nukleinsavat a baktériumba. - az injekció során a fág kapszidja kívül marad, ebben az esetben a penetráció és a dekapszidáció egyetlen lépésben zajlik. - a behatolás a baktérium piluson keresztül is megvalósulhat a konjugáció során. 3. Dekapszidáció a sejtbe jutott vírusrészecskéből kiszabadul a nukleinsav - elveszítik a burkot vagy a kapszidot a sejtbe való bejutáskor - a kapszid eljut a nukleáris pórusokhoz, kiszabadul a DNS. - a kapszidfehérjéket a sejt saját enzimjei, főleg a lizoszómákból származók emésztik le. - a Poxvírusok maguk is kódolnak dekapszidációt befejező litikus enzimeket. - a vírusreplikáció eddigi lépéseinek csekély a hatékonysága. Az
adszorbeálódott vírusok egy része nem is jut a sejtbe, a bejutó genomok enzimatikusan károsodhatnak. 4. Eklipsz fázis - a vírusszaporodás szintetikus szakasza; - sötét szakasz, a virion a sejtben semmiféle módszerrel nem mutatható ki; - dekapszidációtól az új vírusrészecskék megjelenéséig tart; - vírusfehérjék szintézise a gazdasejt fehérjeszintetizáló rendszerével; - a vírusokra jellemző a genom sejten belüli szaporodásának helye. - az eklipszis korai szakaszában a vírusnukleinsav megfelelő szekvenciái átíródnak mRNS-re – transzkripció. - a transzkripció által átírt mRNS transzlációja a gazdasejt riboszómáin történik, aminek eredményeképpen megjelennek az első vírusspecifikus fehérjék a korai fehérjék. - a korai fehérjék nem épülnek be a később képződő új virionokba. Működésük szerint egyrészt olyan enzimek amelyek a vírus szaporodásához a továbbiakban feltétlenül szükségesek (pl.
nukleinsav-polimerázok a vírus nukleinsavának replikációját segítik), másrészt pedig represszorok (olyan fehérjék, amelyek elnyomják a sejt eredeti anyagcsere-tevékenységét). - a korai fehérjék megjelenésével az eklipsz fázis korai szakasza befejeződik, és megkezdődik az új virion nukleinsavának másolata az eredeti nukleinsavról, a korai fehérjék között szereplő polimerázok közreműködésével a vírus nukleinsav replikációja - a következő lépés az új vírusnukleinsavak transzkripciója mRNS-re és a sejtriboszómákon a késői fehérjék szintézise. - a késői fehérjék olyan vírusspecifikus molekulák, amelyek az újonnan képződő virionoknak lesznek alkotórészei: kapszid és belső fehérjék. 5. Morfogenézis - az újonnan szintetizálódott vírusgenomok és struktúrfehérjék komplett vírusrészecskévé épülnek össze. - a vírusérés (maturáció) során konformációs, strukturális változások történnek a
kapszid-fehérjékben, a nukleoproteinek a genommal kondenzálódnak. A vírus ebben a lépésben válik fertőzővé. - vírusérés a peplonnal rendelkező vírusoknál csak a sejtmembránon való kitüremkedéssel ér véget. 6. Kiszabadulás - rendszerint a fertőzött sejtek szétesésével jár, de sok esetben a sejtek csak fokozatosan pusztulnak el, vagy akár hosszú ideig túl élhetik a fertőzést. - a vírus érési ideje és a sejtből való kiszabadulás kezdete a fertőzéstől számítva általában eltérő a különböző vírusoknál és jellemző az egyes családokra. - egy-egy fertőzött sejtben a körülményektől és a vírustól függően több ezer, több tízezer, esetleg százezres nagyságrendű új virion is képződhet, a virionná össze nem épülő, feleslegben termelt vírusalkatrészek mellett. Vírusinterferencia - az a jelenség amikor valamely vírus bejutása a sejtbe megakadályozza ugyanennek a sejtnek egy másik vírussal való
fertőzését. Ez lehetséges: - ugyanazon vírus különböző változatai, - ugyanazon vírus élő és inaktivált formái, - különböző vírusfajták között. Több módon is kialakulhat: ha 2 vírusnak hasonlóak a sejtfelszíni receptorai, egymás kötődését akadályozhatják (pl. retrovírusok, enterovírusok); az adszorbciót a receptor elbontásával teszik hatástalanná (pl. orthomyxovírus); az egyik vírus gátolhatja a másik valamely fontos enzimjét; az első vírusfertőzés hatására a sejt inhibitort is termelhet. - autointerferencia: a vírus saját szaporodását akadályozza. Ebben az esetben a komplett virionok helyett defektív interferáló víruspartikulák képződnek, amelyek önmagukban szaporodni képtelenek. - számos vírusnál előfordul (pl. influenza vírus) és főleg abban az esetben ha jelentős vírusmennyiséggel történik a fertőzés. Vírusexaltáció: két vírus egymásnak kölcsönösen kedvezőbb
feltételeket teremthet a sejtbe való bejutásra és szaporodásra. A vírusok genetikája Mutáció Rekombináció Fenotípusos és genotípusos keveredés Komplementáció és interdependencia 1. Mutáció - a vírusnukleinsavban történő változás, a vírusgén által kódolt fehérje megváltozását jelenti. Spontán mutációk Indukált mutációk sugárzással kémiai mutagénekkel: in vitro reagálnak a vírusnukleinsavval vagy a sejtben replikálódó vírusra hatnak. Lehetnek: - bázisanalógok - módosíthatják a bázisok kémiai szerkezetét - eltávolíthatnak egyes bázisokat - új bázist inszertálhatnak - a molekuláris biológia újabb módszereivel célzott mutációk is kialakíthatók pl. restrikciós fragmentdeléciós módszer, inszerciós, szubsztituciós módszerek. - ha stabil vírusanyagra van szükség (pl. oltóanyaggyártáshoz) akkor a tenyészet genetikai homogenitása egyetlen plakk vagyis egyetlen fertőző virionból kiindulva
biztosítható úgy, hogy ellenőrzött körülmények között szaporítjuk. Vad-típusú vírus az eredeti vírus amelyből a mutánsok származnak, amivel a mutánsokat összehasonlítjuk. Primer izolátum a természetes gazdaszervezetből újonnan izolált vírus. Defektív vírus amikor a vírusreplikációhoz szükséges egy vagy több funkcionális gén hiányzik a virionból. - a mutánsok felismerését az teszi lehetővé, hogy valamely fenotípusos tulajdonságuk eltérő a vad típusétól. - a mutáció a genotípusban beállott változás, esetenként a fenotípus megváltozásában is kifejezésre jut. Plakk-típus mutáció - megfelelő kísérleti körülmények között a vírusok a sejttenyészeteken nekrotikus foltokat – plakkokat hoznak létre, melyek átmérője, külleme nagyjából azonos. A mutáns által létrehozott plakk nagysága vagy egyéb tulajdonsága eltérő, ennek alapján a mutáns izolálható és tanulmányozható. 2.
Rekombináció két különböző, de rokon vírus nukleinsavának kisebbnagyobb szegmentumai kicserélődnek, a keletkező hibrid vírusok mindkét elődtől származó, új génkombinációt tartalmaznak. Előfordul: - kettősláncú DNS vírusok esetén - szegmentált genomot tartalmazó vírusok esetén Létrejöhet nem rokon vírusok között is, vagy a sejt és a vírus DNS között Klaszikus rekombináció Keresztreaktiválás: aktív (infektív) vírusba önmagában szaporodásra képtelen inaktivált vírus aktívan maradt génjei épülnek be. Többszörösségi reaktiválás: infektív virionok képződnek olyan sejtben, amelyet ugyanazon vírustörzsből származó két vagy több különböző letális mutánssal fertőztek. Így épen maradt gének rekombinációjának eredményeként infektív virionok tudnak kialakulni. 3. Fenotípusos és genotípusos keveredés Fenotípusos keveredés: közös tulajdonságokkal rendelkező vírusokkal történt kettős
fertőzés során az utódvírusok mindkét előd fenotípusos tulajdonságait megszerzik, de genotípusuk érintetlen marad. - peplonnal rendelkező vírusoknál az utódvírusok peplonja mindkét vírusra jellemző antigéneket tartalmazza. A virionban csak az egyik szülői nukleinsav van jelen. Nincs változás a genomban Gazdaspektrum változása Rezisztencia a semlegesítő ellenanyagokkal szemben - peplonnal nem rendelkező vírusoknál a fenotípusos keveredést a transzkapszidáció jelensége mutatja. Ez a kapszidnak teljes vagy részleges cseréjét jelenti. genotípusos keveredés nem stabil genetikai változás, két különböző, komplett vírusgenom épül be egyetlen kapszidba, véletlenszerűen. - egy adott víruspartikulából kétféle utódgeneráció származik. 4. Komplementáció és interdependencia - ha kettősen vagy többszörösen fertőzött sejttenyészetekben a különböző vírusok „génproduktumai”: fehérjék, enzimek olyan
kölcsönhatásba lépnek, hogy az elősegíti az egyik vagy mindegyik vírus szaporodását komplementációról beszélünk. - nincs nukleinsavcsere, hanem az egyik vírus olyan enzimet vagy más fehérjét kódol, amelyet a másik nem, de amely szükséges az utóbbi szaporodásához. Interdependencia egyes vírusok egyoldalú vagy kölcsönös függő viszonya. - egyes vírusok esetében szaporodásukhoz még egy másik vírus jelenléte is szükséges a sejtben, függenek a másik vírustól – szatellitái más vírusoknak. A „helper”, „segítő” vírus ellenszolgáltatás nélkül gondoskodik egy enzimről vagy enzimekről, amelyek a szaporodáshoz szükségesek. A környezeti tényezők hatása a vírusokra Fizikai tényezők 1. Hőmérséklet - a vírusok általában csak olyan hőmérsékleti értékeknél szaporodnak amelyek azonosak vagy hasonlóak a gazdaszervezet optimális hőmérsékletével. - a hideggel szemben a vírusok ellenállók, főleg
liofilizált állapotban. 2. Nedvesség - kiszáradva a legtöbb vírus elpusztul. 3. Sugárzások - az ultraibolya és más sugarak elpusztítják a vírusokat. 2. Kémiai tényezők - különféle vegyi anyagok (fenol, savak és bázisok, formaldehid), fertőtlenítőszerek megfelelő mennyiségben inaktiválják és elpusztítják a vírusokat. - antitestek, interferon. A vírusok tropizmusa - az állatpatogén vírusokra vonatkozóan vezették be. - affinitást jelent bizonyos állatfajokhoz genotropizmus szövetekhez hisztotropizmus sejtekhez citotropizmus hisztotropizmus: - dermotropimus (pl. himlővírus) - neurotropizmus (pl. a veszetség vírusa) - pneumotropizmus (pl. grippa vírus) - viszcerotropimus (pl. a sárga láz vírusa) - pantropimus (pl. sertéspestis vírus) A vírusok okozta kóros elváltozások Növénypatogén vírusok klorózis, nekrózis (szövetelhalás), a levelek különféle deformációi (ráncosodás, atrófia), a levelek
összesodrodása, zárványtestek képződése a sejtekben, antocián szintézis gátlás. Állatpatogén vírusok citopatogén hatás daganatkeltő hatás zárványtestképző hatás gyulladást kiváltó hatás - a sejtbe bejutott vírus olyan változásokat vált ki amelyek fénymikroszkóppal is megfigyelhetők: - a sejtek összehúzódnak, legömbölyödnek, a citoplazma szemcsés szerkezetű lesz; - a sejtek megnőnek, legömbölyödnek és számos sejt csoportot képezve összetapad; - a citoplazmában vakuólák képződnek; - egyes sejtek soksejtmagvú óriássejtekké alakulhatnak át; - a sejtek elpusztulnak (citicid hatás) és lizálódnak (citolitikus hatás) tarfoltok képződnek. Bakteriofágok a prokarióták – baktériumok vírusai. - a baktériumokban elszaporodva feoldják azokat (phageo=elfogyasztani). - a folyékony táptalaj feltisztul, szilárd táptalajon a baktériumtenyészeten tarfoltok jelennek meg. - a bakteriofágok nukleinsava lehet DNS
vagy RNS, egyszálú vagy kétszálú. - egyes bakteriofágok külső burokkal peplonnal is rendelkeznek, amelyben jelentős mennyiségű lipid található. - tudományos jelentőségük nagy. - leginkább tanulmányozott fágok az E. Coli T-sorozatú ( T2, T4, T6) farkos fágjai. Kétszállú DNS-t tartalmaznak amely a fág kubikális fejében található és amelyhez kontraktilis farok csatlakozik. A farok legvégén található alaplemezből nyúlványok eredhetnek. - szaporodásmódjuk szerint a fágok lehetnek: 1. litikus fágok 2. temperált fágok - a litikus fágok a szaporodásuk során nagyszámú új fágot produkálva megölik, feloldják a gazdasejtet. - a temperált fágok nem minden esetben okozzák a gazdabaktérium pusztulását, hanem nem-litikus profág formájában is létezhetnek. A fág nukleinsava beépül a gazdasejt nukleinsavába. - a profágot hordozó baktériumsejtekben a fág replikációját egy represszor jellegű fehérje gátolja lizogén
állapot. - a fággenom magával vihet kromoszómadarabokat a gazdasejtből, vagy fág-DNS rész maradhat a baktériumba, a gazdasejt egyes tulajdonságainak megváltozását okozva. pl. toxint nem termelő baktériumok toxintermelővé válhatnak, megváltozhat egyes antibiotikumokra való érzékenységük lizogén konverzió. - transzdukció bakteriofágok által közvetített nukleinsav átvitel. - egy bizonyos fág általában csak 1 baktériumfajt fertőz szigorú gazdaspecificitás
hozzáférhetőségét. - a baktériumok fágkötő receptorai a sejtfalban találhatók. - a Gram negatív baktériumok esetében lipopoliszaharid természetűek, a Gram pozitív baktériumoknál mureinhez kötött teichoinsavakból állnak. - a fágok a farok rostjaival és alaplapjával, illetve poliszaharidáz enzim segítségével valósítják meg a kötődést a baktériumsejt receptoraihoz. 2. Bejutás - penetráció - a növénypatogén vírusok esetében a teljes virion hatol be a növényi gazdasejtbe. A sejtfalak mechanikai sérülését követi, amit az ízeltlábúak szúrásai, jégverés, növényápolási munkálatok okoznak. - a humán- és állatpatogén vírusok behatolása 2 mechanizmus révén valósul meg: endocitózis sejtfúzió - endocitózis során a gazdasejt membránja betűremkedik és bekebelezi a viriont mely egy vakuólába jut. A vírus bekebelezése után helyreáll a sejthártya épsége. - a penetrációnak ezt a formáját
viropexisnek is nevezik. - fúzió során a vírusburok (peplon) fuzionál a sejtmembránnal, egy csatorna képződik amelyen át a vírus bejut a citoplazmába. - a bakterifágok esetében a kontraktilis fehérjetok összehúzódik, a fág belöveli a nukleinsavat a baktériumba. - az injekció során a fág kapszidja kívül marad, ebben az esetben a penetráció és a dekapszidáció egyetlen lépésben zajlik. - a behatolás a baktérium piluson keresztül is megvalósulhat a konjugáció során. 3. Dekapszidáció a sejtbe jutott vírusrészecskéből kiszabadul a nukleinsav - elveszítik a burkot vagy a kapszidot a sejtbe való bejutáskor - a kapszid eljut a nukleáris pórusokhoz, kiszabadul a DNS. - a kapszidfehérjéket a sejt saját enzimjei, főleg a lizoszómákból származók emésztik le. - a Poxvírusok maguk is kódolnak dekapszidációt befejező litikus enzimeket. - a vírusreplikáció eddigi lépéseinek csekély a hatékonysága. Az
adszorbeálódott vírusok egy része nem is jut a sejtbe, a bejutó genomok enzimatikusan károsodhatnak. 4. Eklipsz fázis - a vírusszaporodás szintetikus szakasza; - sötét szakasz, a virion a sejtben semmiféle módszerrel nem mutatható ki; - dekapszidációtól az új vírusrészecskék megjelenéséig tart; - vírusfehérjék szintézise a gazdasejt fehérjeszintetizáló rendszerével; - a vírusokra jellemző a genom sejten belüli szaporodásának helye. - az eklipszis korai szakaszában a vírusnukleinsav megfelelő szekvenciái átíródnak mRNS-re – transzkripció. - a transzkripció által átírt mRNS transzlációja a gazdasejt riboszómáin történik, aminek eredményeképpen megjelennek az első vírusspecifikus fehérjék a korai fehérjék. - a korai fehérjék nem épülnek be a később képződő új virionokba. Működésük szerint egyrészt olyan enzimek amelyek a vírus szaporodásához a továbbiakban feltétlenül szükségesek (pl.
nukleinsav-polimerázok a vírus nukleinsavának replikációját segítik), másrészt pedig represszorok (olyan fehérjék, amelyek elnyomják a sejt eredeti anyagcsere-tevékenységét). - a korai fehérjék megjelenésével az eklipsz fázis korai szakasza befejeződik, és megkezdődik az új virion nukleinsavának másolata az eredeti nukleinsavról, a korai fehérjék között szereplő polimerázok közreműködésével a vírus nukleinsav replikációja - a következő lépés az új vírusnukleinsavak transzkripciója mRNS-re és a sejtriboszómákon a késői fehérjék szintézise. - a késői fehérjék olyan vírusspecifikus molekulák, amelyek az újonnan képződő virionoknak lesznek alkotórészei: kapszid és belső fehérjék. 5. Morfogenézis - az újonnan szintetizálódott vírusgenomok és struktúrfehérjék komplett vírusrészecskévé épülnek össze. - a vírusérés (maturáció) során konformációs, strukturális változások történnek a
kapszid-fehérjékben, a nukleoproteinek a genommal kondenzálódnak. A vírus ebben a lépésben válik fertőzővé. - vírusérés a peplonnal rendelkező vírusoknál csak a sejtmembránon való kitüremkedéssel ér véget. 6. Kiszabadulás - rendszerint a fertőzött sejtek szétesésével jár, de sok esetben a sejtek csak fokozatosan pusztulnak el, vagy akár hosszú ideig túl élhetik a fertőzést. - a vírus érési ideje és a sejtből való kiszabadulás kezdete a fertőzéstől számítva általában eltérő a különböző vírusoknál és jellemző az egyes családokra. - egy-egy fertőzött sejtben a körülményektől és a vírustól függően több ezer, több tízezer, esetleg százezres nagyságrendű új virion is képződhet, a virionná össze nem épülő, feleslegben termelt vírusalkatrészek mellett. Vírusinterferencia - az a jelenség amikor valamely vírus bejutása a sejtbe megakadályozza ugyanennek a sejtnek egy másik vírussal való
fertőzését. Ez lehetséges: - ugyanazon vírus különböző változatai, - ugyanazon vírus élő és inaktivált formái, - különböző vírusfajták között. Több módon is kialakulhat: ha 2 vírusnak hasonlóak a sejtfelszíni receptorai, egymás kötődését akadályozhatják (pl. retrovírusok, enterovírusok); az adszorbciót a receptor elbontásával teszik hatástalanná (pl. orthomyxovírus); az egyik vírus gátolhatja a másik valamely fontos enzimjét; az első vírusfertőzés hatására a sejt inhibitort is termelhet. - autointerferencia: a vírus saját szaporodását akadályozza. Ebben az esetben a komplett virionok helyett defektív interferáló víruspartikulák képződnek, amelyek önmagukban szaporodni képtelenek. - számos vírusnál előfordul (pl. influenza vírus) és főleg abban az esetben ha jelentős vírusmennyiséggel történik a fertőzés. Vírusexaltáció: két vírus egymásnak kölcsönösen kedvezőbb
feltételeket teremthet a sejtbe való bejutásra és szaporodásra. A vírusok genetikája Mutáció Rekombináció Fenotípusos és genotípusos keveredés Komplementáció és interdependencia 1. Mutáció - a vírusnukleinsavban történő változás, a vírusgén által kódolt fehérje megváltozását jelenti. Spontán mutációk Indukált mutációk sugárzással kémiai mutagénekkel: in vitro reagálnak a vírusnukleinsavval vagy a sejtben replikálódó vírusra hatnak. Lehetnek: - bázisanalógok - módosíthatják a bázisok kémiai szerkezetét - eltávolíthatnak egyes bázisokat - új bázist inszertálhatnak - a molekuláris biológia újabb módszereivel célzott mutációk is kialakíthatók pl. restrikciós fragmentdeléciós módszer, inszerciós, szubsztituciós módszerek. - ha stabil vírusanyagra van szükség (pl. oltóanyaggyártáshoz) akkor a tenyészet genetikai homogenitása egyetlen plakk vagyis egyetlen fertőző virionból kiindulva
biztosítható úgy, hogy ellenőrzött körülmények között szaporítjuk. Vad-típusú vírus az eredeti vírus amelyből a mutánsok származnak, amivel a mutánsokat összehasonlítjuk. Primer izolátum a természetes gazdaszervezetből újonnan izolált vírus. Defektív vírus amikor a vírusreplikációhoz szükséges egy vagy több funkcionális gén hiányzik a virionból. - a mutánsok felismerését az teszi lehetővé, hogy valamely fenotípusos tulajdonságuk eltérő a vad típusétól. - a mutáció a genotípusban beállott változás, esetenként a fenotípus megváltozásában is kifejezésre jut. Plakk-típus mutáció - megfelelő kísérleti körülmények között a vírusok a sejttenyészeteken nekrotikus foltokat – plakkokat hoznak létre, melyek átmérője, külleme nagyjából azonos. A mutáns által létrehozott plakk nagysága vagy egyéb tulajdonsága eltérő, ennek alapján a mutáns izolálható és tanulmányozható. 2.
Rekombináció két különböző, de rokon vírus nukleinsavának kisebbnagyobb szegmentumai kicserélődnek, a keletkező hibrid vírusok mindkét elődtől származó, új génkombinációt tartalmaznak. Előfordul: - kettősláncú DNS vírusok esetén - szegmentált genomot tartalmazó vírusok esetén Létrejöhet nem rokon vírusok között is, vagy a sejt és a vírus DNS között Klaszikus rekombináció Keresztreaktiválás: aktív (infektív) vírusba önmagában szaporodásra képtelen inaktivált vírus aktívan maradt génjei épülnek be. Többszörösségi reaktiválás: infektív virionok képződnek olyan sejtben, amelyet ugyanazon vírustörzsből származó két vagy több különböző letális mutánssal fertőztek. Így épen maradt gének rekombinációjának eredményeként infektív virionok tudnak kialakulni. 3. Fenotípusos és genotípusos keveredés Fenotípusos keveredés: közös tulajdonságokkal rendelkező vírusokkal történt kettős
fertőzés során az utódvírusok mindkét előd fenotípusos tulajdonságait megszerzik, de genotípusuk érintetlen marad. - peplonnal rendelkező vírusoknál az utódvírusok peplonja mindkét vírusra jellemző antigéneket tartalmazza. A virionban csak az egyik szülői nukleinsav van jelen. Nincs változás a genomban Gazdaspektrum változása Rezisztencia a semlegesítő ellenanyagokkal szemben - peplonnal nem rendelkező vírusoknál a fenotípusos keveredést a transzkapszidáció jelensége mutatja. Ez a kapszidnak teljes vagy részleges cseréjét jelenti. genotípusos keveredés nem stabil genetikai változás, két különböző, komplett vírusgenom épül be egyetlen kapszidba, véletlenszerűen. - egy adott víruspartikulából kétféle utódgeneráció származik. 4. Komplementáció és interdependencia - ha kettősen vagy többszörösen fertőzött sejttenyészetekben a különböző vírusok „génproduktumai”: fehérjék, enzimek olyan
kölcsönhatásba lépnek, hogy az elősegíti az egyik vagy mindegyik vírus szaporodását komplementációról beszélünk. - nincs nukleinsavcsere, hanem az egyik vírus olyan enzimet vagy más fehérjét kódol, amelyet a másik nem, de amely szükséges az utóbbi szaporodásához. Interdependencia egyes vírusok egyoldalú vagy kölcsönös függő viszonya. - egyes vírusok esetében szaporodásukhoz még egy másik vírus jelenléte is szükséges a sejtben, függenek a másik vírustól – szatellitái más vírusoknak. A „helper”, „segítő” vírus ellenszolgáltatás nélkül gondoskodik egy enzimről vagy enzimekről, amelyek a szaporodáshoz szükségesek. A környezeti tényezők hatása a vírusokra Fizikai tényezők 1. Hőmérséklet - a vírusok általában csak olyan hőmérsékleti értékeknél szaporodnak amelyek azonosak vagy hasonlóak a gazdaszervezet optimális hőmérsékletével. - a hideggel szemben a vírusok ellenállók, főleg
liofilizált állapotban. 2. Nedvesség - kiszáradva a legtöbb vírus elpusztul. 3. Sugárzások - az ultraibolya és más sugarak elpusztítják a vírusokat. 2. Kémiai tényezők - különféle vegyi anyagok (fenol, savak és bázisok, formaldehid), fertőtlenítőszerek megfelelő mennyiségben inaktiválják és elpusztítják a vírusokat. - antitestek, interferon. A vírusok tropizmusa - az állatpatogén vírusokra vonatkozóan vezették be. - affinitást jelent bizonyos állatfajokhoz genotropizmus szövetekhez hisztotropizmus sejtekhez citotropizmus hisztotropizmus: - dermotropimus (pl. himlővírus) - neurotropizmus (pl. a veszetség vírusa) - pneumotropizmus (pl. grippa vírus) - viszcerotropimus (pl. a sárga láz vírusa) - pantropimus (pl. sertéspestis vírus) A vírusok okozta kóros elváltozások Növénypatogén vírusok klorózis, nekrózis (szövetelhalás), a levelek különféle deformációi (ráncosodás, atrófia), a levelek
összesodrodása, zárványtestek képződése a sejtekben, antocián szintézis gátlás. Állatpatogén vírusok citopatogén hatás daganatkeltő hatás zárványtestképző hatás gyulladást kiváltó hatás - a sejtbe bejutott vírus olyan változásokat vált ki amelyek fénymikroszkóppal is megfigyelhetők: - a sejtek összehúzódnak, legömbölyödnek, a citoplazma szemcsés szerkezetű lesz; - a sejtek megnőnek, legömbölyödnek és számos sejt csoportot képezve összetapad; - a citoplazmában vakuólák képződnek; - egyes sejtek soksejtmagvú óriássejtekké alakulhatnak át; - a sejtek elpusztulnak (citicid hatás) és lizálódnak (citolitikus hatás) tarfoltok képződnek. Bakteriofágok a prokarióták – baktériumok vírusai. - a baktériumokban elszaporodva feoldják azokat (phageo=elfogyasztani). - a folyékony táptalaj feltisztul, szilárd táptalajon a baktériumtenyészeten tarfoltok jelennek meg. - a bakteriofágok nukleinsava lehet DNS
vagy RNS, egyszálú vagy kétszálú. - egyes bakteriofágok külső burokkal peplonnal is rendelkeznek, amelyben jelentős mennyiségű lipid található. - tudományos jelentőségük nagy. - leginkább tanulmányozott fágok az E. Coli T-sorozatú ( T2, T4, T6) farkos fágjai. Kétszállú DNS-t tartalmaznak amely a fág kubikális fejében található és amelyhez kontraktilis farok csatlakozik. A farok legvégén található alaplemezből nyúlványok eredhetnek. - szaporodásmódjuk szerint a fágok lehetnek: 1. litikus fágok 2. temperált fágok - a litikus fágok a szaporodásuk során nagyszámú új fágot produkálva megölik, feloldják a gazdasejtet. - a temperált fágok nem minden esetben okozzák a gazdabaktérium pusztulását, hanem nem-litikus profág formájában is létezhetnek. A fág nukleinsava beépül a gazdasejt nukleinsavába. - a profágot hordozó baktériumsejtekben a fág replikációját egy represszor jellegű fehérje gátolja lizogén
állapot. - a fággenom magával vihet kromoszómadarabokat a gazdasejtből, vagy fág-DNS rész maradhat a baktériumba, a gazdasejt egyes tulajdonságainak megváltozását okozva. pl. toxint nem termelő baktériumok toxintermelővé válhatnak, megváltozhat egyes antibiotikumokra való érzékenységük lizogén konverzió. - transzdukció bakteriofágok által közvetített nukleinsav átvitel. - egy bizonyos fág általában csak 1 baktériumfajt fertőz szigorú gazdaspecificitás
 Budapesten született 1887. június 25-én. Apja Karinthy József tisztviselő, anyja Engel Karolina; Frigyes a család ötödik gyermeke.1893-ban meghal édesanyja. Anyjának korai elvesztése, az eleven szellemi környezet és Budapest világvárossá fejlődése meghatározó gyermekkori élménye. Testvérei, Etelka és Elza festőnövendékek, Emília rendkívüli nyelvtehetség. Frigyes - a
Budapesten született 1887. június 25-én. Apja Karinthy József tisztviselő, anyja Engel Karolina; Frigyes a család ötödik gyermeke.1893-ban meghal édesanyja. Anyjának korai elvesztése, az eleven szellemi környezet és Budapest világvárossá fejlődése meghatározó gyermekkori élménye. Testvérei, Etelka és Elza festőnövendékek, Emília rendkívüli nyelvtehetség. Frigyes - a